by Jack D. Mount¹
Introduction
The Phylum Brachiopoda, marine invertebrate animals with a pair of valves resembling the Molluscan
bivalves, can easily be recognized by its bilateral symmetry--one half of the shell is the mirror image of the other half when seen from
the front with the beaks vertical. The brachiopods are divided into two groups by the nature of their shell. The Class Inarticulata is
characterized by a phosphatic shell with the valves held together only with muscles. The other group, the Class Articulata, has
calcareous shells and the valves are fastened together by teeth and sockets.
While both groups first
appear in the Early Cambrian, the articulated brachiopods are uncommon in Cambrian rocks anywhere. Thus, two species of articulate
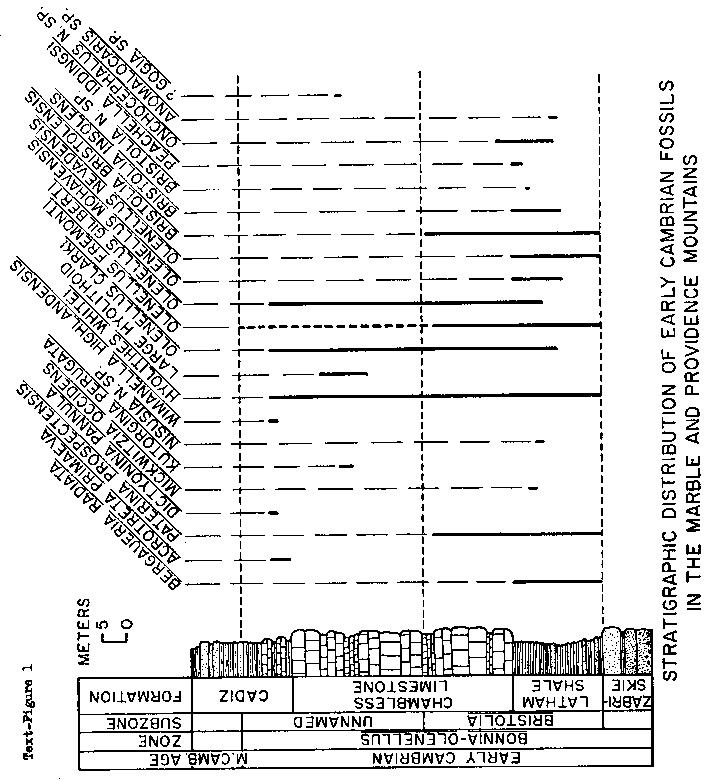
brachippods recovered during my study (Mount, 1973,1974a,1974b) of the highly fossiliferous Lower Cambrian section (see Text-figure 1)
in the Marble Mountains, San Bernardino County, California, provided a pleasant surprise. It is important to note that the
identifications were made with some difficulty and should be considered tentative. The specimens occur in shale and the shells are
missing with only impressions of the interior surface preserved. In modern brachiopod systematics internal morphology is now more
important than shell surface characters.
The following abbreviations are used:
UCR - Department of
Earth Sciences, University of California, Riverside.
USNM - National Museum of Natural History, Washington, D.C.
Order Orthida
Superfamily Billingsellacea
Family Nisusiidae
Genus Nisusia Walcott
Nisusia n.sp.?
Plate 1, figures 1-2
Description of Pedicle Valve: Shell medium in size for the genus; apparently thin; outline transversely subrectangular; wider than long with the greatest width at the hinge; cardinal extremities nearly rectangular; lateral margins nearly straight or slightly convex; anterior margin broadly rounded. Sulcus very shallow and wide, originating in anterior third of shell and extending to anterior margin. Umbo short, widely convex, moderately swollen and protruded posterior to the posterior margin; apical angle 112°. Interarea narrow and apparently apsacline. Ornamentation apparently consists of somewhat irregularly arranged narrowly rounded costae separated by interspaes with widths from 1 to 3 times the width of the costae. Surface marked by concentric lines of growth.
Hypotype: UCR 10/2019 (pl.1, figs.l,2) (collected by John Kniffen); length 11.3 mm, width 15.0 mm; UCR locality 10.
Stratigraphic Position: in gray-green shale in upper half of the Latham Shale of Hazzard (1954).
Age: The following taxa have been collected with the brachiopod described here:
BRACHIOPODA: Paterina pospectensis (Walcott), Mickwitzia occidens Walcott, ?Obolus
sp. indet.
MOLLUSCA: Hyolithes whitei Resser
TRILOBITA: Olenellus clarki (Reeser), 0. fremonti Walcott, O. gilberti Meek,
0. mohavensis (Crickmay), O. nevadensis (Walcott), Bristolia bristolensis (Resser), B. insolens (Resser),
B. n.sp., Onchocephalus n.sp.
MALACOSTRACA: Anomalocaris sp.
This fauna is characteristic of the late Early Cambrian Bonnia-0lenellus Zone of Rasetti (1951),
the youngest of the three Early Cambrian zones (Fritz, 1972), and is assigned to the "Bristolia Subsone" of Mount (1974,1974b).
Discussion: This species is not quite like any other described, but because it is only known from one specimen, flattened in shale and shorn of its shell, it is impossible to make detailed comparisons with other better-preserved species, Consequently, the species can not be named until more and better preserved specimens are recovered. The new species appears, however, to be closely related to Nisusia bivia (Walcott)(1912) from the Lower Cambrian Carrara Formation in the Resting Springs Range, Inyo County, California, but differs in having narrower costae with wider interspaces and in other details of surface sculpture.
Wimanella highlandensis (Walcott)
Plate 1, figures 3-8
Orthis? highlandensis Walcott, 1886, p.119, pl.8, figs.3,3a,3b. Walcott, 1891, p.612, pl.72, figs-5,5a,5b. Pack, 1906,
p.292.
Billingsella highlandensis (Walcott), Walcott, 1905, p.237. Walcott, 1912, p.756, pl. 87, figs.4,4a,4b,4c. ?Mason, 1935, p.102.
?Hazzard and Mason, 1936, p.233. Mount, 1974a, p.2, fig.7. Mount, 1974b, p.224.
Wimanella highlandensis (Walcott), Schuchert and Cooper, 1932, p.50. Bell, 1941, p.
240, pl.33, figs.23,24,25.
Holotype: USNM 15355c.
Type Locality: USNM 31a; east side of anticline near Pioche, Lincoln County, Nevada; Pioche Shale, Lower Cambrian.
Original Description: "Shell about the average size of the Cambrian species, but below the size of those from the Silurian.
Transversely oval or subrotund, front broadly rounded and nearly straight in the dorsal valve; hinge line shorter than the greatest
width of the shell, Ventral valve moderately convex, most elevated toward the beak, which is slightly arched over to meet the nearly
perpendicular area; mesial sinus broad and shallow or nearly obsolete; area flat; foramen unknown. Dorsal valve associated in the same
hard specimens of limestone of limestone more convex than the ventral valve; hinge line very short; median fold but slightly raised
above the general surface; area unknown. Surface marked by concentric lines of growth and finer striae. No traces of radiating costae
are seen except on the interior of the shell towards the margin.
Casts of the interior of the ventral
valve show the dental plates, median ridge and bifurcating ridges extending nearly to the margin; also two lateral grooves. The
specimens of the interior of the dorsal valve show only faint impressions of a large oval scar near each side of the median line."
(Walcott, 1886, p.119)
Supplementary Description: "Shell transversely oval or subrotund. Front broadly rounded and nearly straight in the dorsal
valve. On the ventral valve the cardinal line slopes toward the beak at an angle of about 20°, while in the dorsal valve it is lose
than 10°. The greatest width is about the middle of the shell. The ventral valve is moderately convex, being most elevated toward
the beak, which is slightly arched over to the cardinal margin.
A dorsal valve associated in the sane
hand specimen of limestone is more convex than the ventral valve. The cardinal line is shorter than the greatest width of the shell.
The median fold is but slightly raised above the general surface, although the front margin has a low, broad arch for the reception of
the fold of the ventral valve.
A slight flattening in the central portion of the ventral valve in
some specimens forms a low, broad menial sinus toward the front margin.
The surface is marked by
concentric lines of growth and fine radiating ribs; on some specimens stronger ribs occur very much as on Billingsella
coloradoensis. Interior casts of the valves show fine, radiating striae toward the front. In a number of partly exfoliated shells,
the shell is shown to be thick and apparently solid, having been replaced by calcite.
The
average-sized ventral valve has a length of 13 mm., with a width of 16mm. One dorsal valve is 17 mm. in width.
Cardinal area of the ventral valve unknown except that its plane extends backward at an angle of about
10° or 15° to the plane of the margin of the shell.
Casts of the interior of the ventral
valve show that the dental plates extend down to the bottom of the valve, supporting distinctly defined hinge teeth. The traces of a
vascular system are limited to the main vascular trunks, which extend forward well toward the front margin, very much as in B.
coloradoensis. In one cast there is a strong furrow extending from a median furrow obliquely outward to each main vascular sinus.
The median furrow extends backward to the apex of the cast that filled the space beneath the umbo and the deltidium. This portion of the
cast is also marked by fine vertical venation. In another cast there is a very narrow median furrow. These median furrows probably
indicate the beginning of a septum that in later forms connected the deltidium with the shell. Nothing is known of the interior of the
dorsal valve." (Walcott, 19129 p.756-757)
Hypotypes: UCR 7307/4 (pl.1, fig-3) (collected by James E. Fuller), length 13.2 mm, width 17.4 mm; UCR 7307/5 (pl.1, fig-4)9 length 16.7 mm, width 19.7 mm; UCR 7307/12 (pl.1, fig-5), length 14.5 mm, width 19.6 mm; UCR 7307/13 (pl.1, figs.6,7), length 10.0 mm, width 14.1 mm; all UCR locality 7307. UCR 7527/1 (pl.1, fig.8); length 13.7 mm, width 19.1 mm; UCR locality 7527.
Stratigraphic Position: UCR localities 7307 and 7527 are in the Cadiz Formation of Hazard and Mason (1936) and Hazzard (1954), in reddish-brown shale 3 meters above the base of the formation.
Age: At UCR localities 7307 and 7527 the following fossils have been collected:
BRACHIOPODA: Acrotreta primaeva Walcott, Dictyonina pannula (White)
MOLLUSCA: Hyolithes whitei Resser
TRILOBITA: Olenellus clarki (Resser), O. gilberti Meek.
These taxa occur in the late Early Cambrian Bonnia-Olenellus Zone and are assigned to the latest,
as yet unnamed, subzone of the Early Cambrian (see Text-figure 1).
Distribution: Pioche Hills and Highland Range, Lincoln County, Nevada; Pioche Shale, Lower Cambrian. Inyo Mountains, Inyo County, California; probably Saline Valley Formation, Lower Cambrian. Marble Mountains, San Bernardino County, California; Cadiz Formation, Lower and ?Middle Cambrian.
Discussion: Wimanella highlandensis is common in the single horizon noted above. In most details of the internal surface the Marble Mountains specimens compare favorably with the type specimens from Nevada. Several of the Cadiz specimens have a more pronounced sulcus than the Nevada forms, while in other specimens the sulcus is not evident. This feature may be due to distortion of the shell during preservation.
UCR 10: opposite side of ridge from old quarry in the southern end of Marble Mountains; 125 m west and 650 m south of northeast corner of section 11, T. 5 N., R. 14 E., Danby 15' quadrangle (1956 edition); same as locality M-5 of Hazzard (1933). Lathan Shale.
UCR 7307: at elevation of 384 m on east slope of first hill north of hill 1645 in southern end of Marble Mountains; 270 m west and 240 m south of the northeast corner of section 11, T. 5 N., R. 14 E., Danby 15' quad. Cadiz Fm.
UCR 7527: at elevation of 372 m in bottom of deep, narrow gully on east flank of hill 1645 in southern end of Marble Mountains; 240 m west and 480 m south of northeast corner of section 11, T. 5 N., R. 14 E., Danby 15' quad. Cadiz Fm.
Bell, W. C., 1941. Cambrian Brachiopoda from Montana. Jour. Paleo. 15:l93-255, pls.28-37.
Fritz, W. H., 1972. Lower Cambrian trilobites from the Sekwi Formation type section, Mackenzie Mountains, northwestern Canada. Geol.
Survey Canada Bull. 212:l-90, 20 pls.
Hazzard, J. C., 1933. Notes on the Cambrian rocks of the eastern Mohave Desert, California. Univ. California, Dept. Geol. Sci.
Bull. 23:7-80, pl.1.
_____, 1954. Rocks and structure of the northern Providence Mountains, San Bernardino County, California. California Div. Mines
Geol. Bull. 170(4):27-35.
_____, and Mason, J. F., 1936. Middle Cambrian formations of the Providence and Marble Mountains, California. Geol. Soc. America
Bull. 47:229-240.
Mason, J. F., 1935. Fauna of the Cambrian Cadiz Formation, Marble Mountains, California. So. California Acad. Sci. Bull.
34:97-119, pl.15.
Mount, J. D., 1973. Early Cambrian fauna of the Latham Shale, southern California.
So. California Acad. Sci. Ann. Mtg., Abstracts of Papers, p.10.
_____, 1974a. Early Cambrian faunas from the Marble and Providence Mountains, San Bernardino County,, California. Bull. So.
California Paleo. Soc. 6:l-5, 23 figs.
_____, 1974b. Biostratigraphy of the Lower Cambrian in southeastern California. Geol. Soc. America, Abstracts With Programs
6(3):224.
Pack, F. J., 1906. Cambrian fossils from the Pioche Mountains, Nevada. Jour. Geol. 14:290-302, 3 pls.
Rasetti, F., 1951. Middle Cambrian stratigraphy and faunas of the Canadian Rocky Mountains. Smithsonian Misc. Colln.
116(5):1-277, 34 pls.
Schuchert, C., and Cooper, G. A., 1932. Brachiopod genera of the Subclass Orthoidea and Pentameroidea. Mem. Peabody Mus. Nat.
Hist. 4(1):1-270, 29 pls.
Walcott, C. D., 1886. Second contribution to the studies on the Cambrian faunas of North America. U. S. Geol. Survey Bull.
30:1-369, 33 pls.
_____, 1891. The fauna of the Lower Cambrian or Olenellus Zone. Tenth Ann. Rept. U. S. Geol. Survey (1):509-774,
pls.49-98.
_____, 1905. Cambrian Brachiopoda, with descriptions of new genera and species. U. S. Nat. Mus. Proc. 28:227-337.
_____, 1912. Cambrian Brachiopoda. U. S. Geol. Survey Mon 51:1-872, 104 pls.
I am grateful to Messrs. John Kniffen and James E. Fuller who collected several of the specimens used in this study and generously donated them to the Department of Earth Sciences, University of California, Riverside.
Explanation of Plate 1
All figures x2
Figs. 1-2. Nisusia n.sp.? 1,2. cast and mold of pedicle valve interior, Hypotype, UCR 10/2019; UCR loc. 10, Latham Shale, Lower Cambrian.
Figs. 3-8. Wimanella highlandensis (Walcott). 3, cast of brachial valve interior, Hypotype, UCR 7307/4; UCR loc. 7307, Cadiz
Formation, Lower Cambrian. 4, mold of brachial valve interior, Hypotype, UCR 7307/5; UCR loc. 7307. 5, cast of brachial valve interior,
Hypotype, UCR 7307/12; UCR loc. 7307. 6, 7, cast and mold of brachial valve interior, Hypotype, UCR 7307/13; UCR loc. 7307. 8, cast of
pedicle valve interior, Hypotype, UCR 7527/1; UCR loc. 7527, Cadiz Formation, Lower Cambrian.

_______________
¹Department of Earth Sciences, University of California, Riverside, California 92502.
![]() Return to
Paleontology Resources page
Return to
Paleontology Resources page
![]() Return to first page
of Jack's Home Place
Return to first page
of Jack's Home Place
|
|
 | |
Fossils & Paleontology NetRing | |