CHARACTERISTICS OF EARLY CAMBRIAN FAUNAS FROM EASTERN SAN BERNARDINO COUNTY, CALIFORNIA
by Jack D. Mount¹
Introduction
Moderately to highly fossiliferous lower Cambrian clastic and carbonate rocks crop out at localities scattered throughout much of the Mojave Desert in eastern San Bernardino County, California. This relatively thin succession of strata is representative of the Craton facies of the Cordilleran geocline and rests on crystalline basement rocks of Early Precambrian age. The original discovery of fossiliferous lower Cambrian rocks was made more than 70 years ago in the southern end of the Marble Mountains about 3.3 kilometers northeast of the railroad siding of Cadiz. Similar sections have since been located in the Providence, Kelso, New York and Mesquite Mountains, the Mescal Range, and the Silurian and Salt Spring Hills, all in eastern San Bernardino County. The present report, which is seventh in a series of preliminary notes making known the results of my research on the faunas from these rocks, lists 35 Early Cambrian fossils, which is four times the number of taxa known prior to the initiation of my studies.
More than 40 formal and informal papers, notes and abstracts have been published dealing with aspects of the paleontology and stratigraphy of the study area; however, only the more useful references are discussed here:
Darton (1907, 1915) first described the lower Cambrian section in the southern Marble Mountains (he called-them the Iron Mountains) as part of his study of the geology along the route of the Santa Fe Railroad. Clark (1921) described in more detail this same section, which he referred to as Bristol Mountain, and listed four trilobite taxa. Using Clark's original collection from the classical locality near the old Vaughan Quarry at the southern end of the Marble Mountains, Resser (1928) prepared the first systematic report on the fossils describing 6 taxa, 3 of them new olenellid trilobites.
As the results of studies for graduate degrees at the University of California, J. C. Hazzard published a series of papers (Hazzard, 1933, 1937a, 1938, 1954; Hazzard and Mason, 1936) describing in some detail the Cambrian stratigraphy and formalizing the nomenclature for the previously unnamed rock units in the area. Crickmay (1933) briefly described the fossils and included the description of a new species of olenellid trilobite.
Riccio (1949, 1952) studied the ontogeny and morphology of the olenellid trilobites from the Marble Mountains but made no new additions to the lower Cambrian faunal list for the area.
Five preliminary notes were published by Mount (1973, 1974a, 1974b, 1974c, 1976) outlining the progress of his current study of the systematics and biostratigraphy of the Early Cambrian faunas. Mount (in press) has described one of the oldest articulate brachiopods.
The systematic description of a finely preserved eocrinoid from the Marble Mountains was presented by Durham (1978).
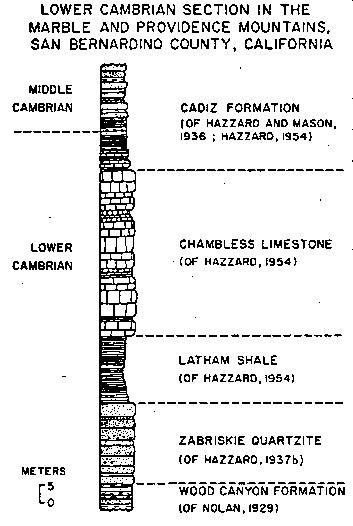
Hazzard (1954) and Stewart (1970) have described the lower Cambrian sections in detail and have divided the lower Cambrian into 5 formations (see text-fig. A). The lower Cambrian rocks consist of a conformable sequence of shallow marine, clastic and carbonate sediments which overlies cratonic Precambrian plutonic or metamorphic rocks to the south and Precambrian marine sediments to the north. The youngest Precambrian basement rock, a porphyritic, coarse-grained granite, in the Marble Mountains area has been dated at 1.4 billion years (Lanphere, 1964). The lowest sedimentary unit in the area, the Wood Canyon Formation of Nolan (1929), consists of from 125 to over 400 meters of a varied assemblage of rocks ranging from shale to sandstone, quartzite and conglomerate. No fossils have been found in it in the study area. Hazzard (1937b) and Cloud (1973) report trilobites from the Wood Canyon Formation in the Nopah Range. Nelson (1976) has shown that the Wood Canyon Formation is entirely of Early Cambrian age and that the Fallotaspis, Nevadella and Bonnia-Olenellus Zones fall with in it.
Massive, poorly fossiliferous, pink to white ortho- and protoquartzite overlie the Wood Canyon Formation. This is the Zabriskie Quartzite of Hazzard (1937b) and it ranges in thickness from less than 5 meters in the Marble Mountains to over 75 meters in the Salt Spring Hills (Stewart, 1970). In some sections these two formations have been lumped together in earlier reports as the Prospect Mountains Quartzite (Hazzard, 1954; Hewett, 1956); the Tapeats Sandstone (Hewett, 1956; Stewart, 1970) or unnamed quartzite (Fritz, 1975).
| Text-fig. A |
The sandstone and quartzite is conformably overlain by the highly fossiliferous Latham Shale of Hazzard (1954), 15 to 45 meters of gray-green shale, platy micaceous siltstone, sandstone and a minor amount of limestone. The shale weathers to red, paper-thin fragments and contains a diverse assemblage of Early Cambrian fossils with abundant olenellid trilobites.
Above the shale is the Chambless Limestone of Hazzard (1954) which ranges from 36 to 66 meters in thickness. It is characterized by the massive beds of algal oncolites of the genus Girvanella (Hazzard, 1937a). Platy limestone with trilobite fragments, brachiopods and primitive mollusks and a minor amount of shale occur as interbeds.
The highest lower Cambrian unit is the Cadiz Formation of Hazzard and Mason (1936). Hazzard (1954) redefined the formation by lowering the lower contact to include the 33+ meters of lower Cambrian rocks excluded in the original definition. The formation consists of a heterogeneous assemblage of rocks including sandstone, shale, siltstone and limestone. Only about the lower one tenth of the formation is Early Cambrian in age. The Cadiz Formation is overlain by the middle Cambrian Bonanza King Formation of Hazzard and Mason (1936).
The fossils thus far recovered from the lower Cambrian rocks total 35 taxa; they are listed in Table 1. Of the 33 taxa identifiable to the species level, 3 are new genera and 12 are new species.
The Latham Shale is dominated by a large quantity and a great diversity of olenellid trilobites of the genera Olenellus, Bristolia and Peachella. Olenellus n. sp. A is characterized by the greatly advanced genal spines and the more acute metagenal angle. The genus Bristolia, which was first recognized from the Latham Shale, is characterized by having very advanced genal spines and an hour-glass shaped glabella that reaches anteriorly to the rim. Bristolia n. sp. has the least advanced genal spines of any species in the genus, a strongly semicircular shaped cephalon and wider palpabral lobes. Onchocephalus n. sp. is represented by 6 known specimens which are the largest and most complete known in the genus. This is the second known occurrence for the inarticulate brachiopod Mickwitzia which was previously known from the lower part of the Bonnia-Olenellus Zone in the Harkless Formation of Esmeralda Co., Nevada. Nisusia "fulleri" Mount, mss, is one of the earliest described species of the genus (Mount, 1974c and in press). The presence of the soft-bodied animals Anomalocaris canadensis and the new annelid (see fig. 20) is particularly interesting and is reminescent of the Burgess Shale fauna. This is the most southwesterly known occurrence of Anomalocaris. Briggs (1979) has interpreted this genus as an appendage of a large as yet unknown arthropod.
Trilobites in the Chambless Limestone are rare and generally poorly preserved. This is the first record of Olenellus puertoblancoensis from the area between its type locality in Sonora, Mexico, and an occurrence in northwestern Canada (Fritz, 1972). Olenellus n. sp. B has a large cephalon with advanced genal spines, a large inflated anterior glabellar lobe and long palpabral lobes extending to the posterior margin. The hyolithoid mollusk Novitatus has previously been known only from eastern Asia. It is interesting to note here that because specimens of some species of trilobites occur in both the Latham Shale and the Chambless Limestone it is possible to compare specimens which may have original relief with those crushed in the shale and Sundberg (1974) has shown that some dimensions of the cephalon may have been increased differentially by as much as 15% through compression.
Fossils are moderately abundant in two shale beds near the base of the Cadiz Formation but are not as diverse as in-the Latham Shale. The fauna from these highest beds of the lower Cambrian in western North America has been poorly known; recently, Palmer and Halley (1979) have described a small assemblage of trilobites from this stratigraphic position in the Carrara Formation. The new olenellid genus (see fig. 32), which appears to be thus far known only from the Cadiz Formation in the Marble Mountains, has mixed characteristics of the genera Bristolia, Peachella and Olenellus.
The stratigraphic ranges of the trilobite taxa suggest that this sequence of rocks belongs to the upper part of the Bonnia-Olenellus Zone of Rasetti (1951), the youngest of the three Early Cambrian zones (Fritz, 1972). The base of this zone is considered to extend to at least 15 meters below the base of the Latham Shale where a single specimen of Olenellus sp. indet. was collected. The Early Cambrian fossils from eastern San Bernardino County are sufficiently abundant and evenly distributed through the section and can be grouped into three faunules (see fig. 33) demonstrating the need for three subdivisions of the upper part of the Bonnia-Olenellus Zone in this area.
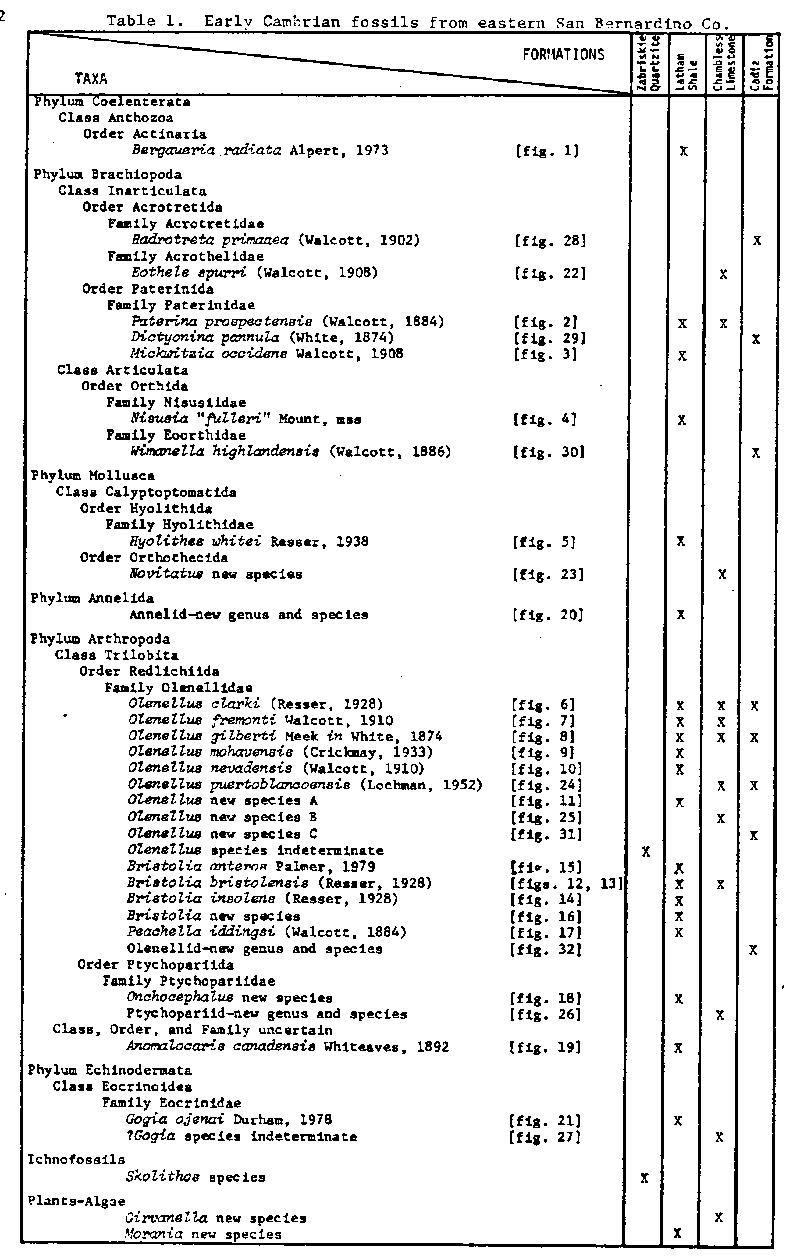
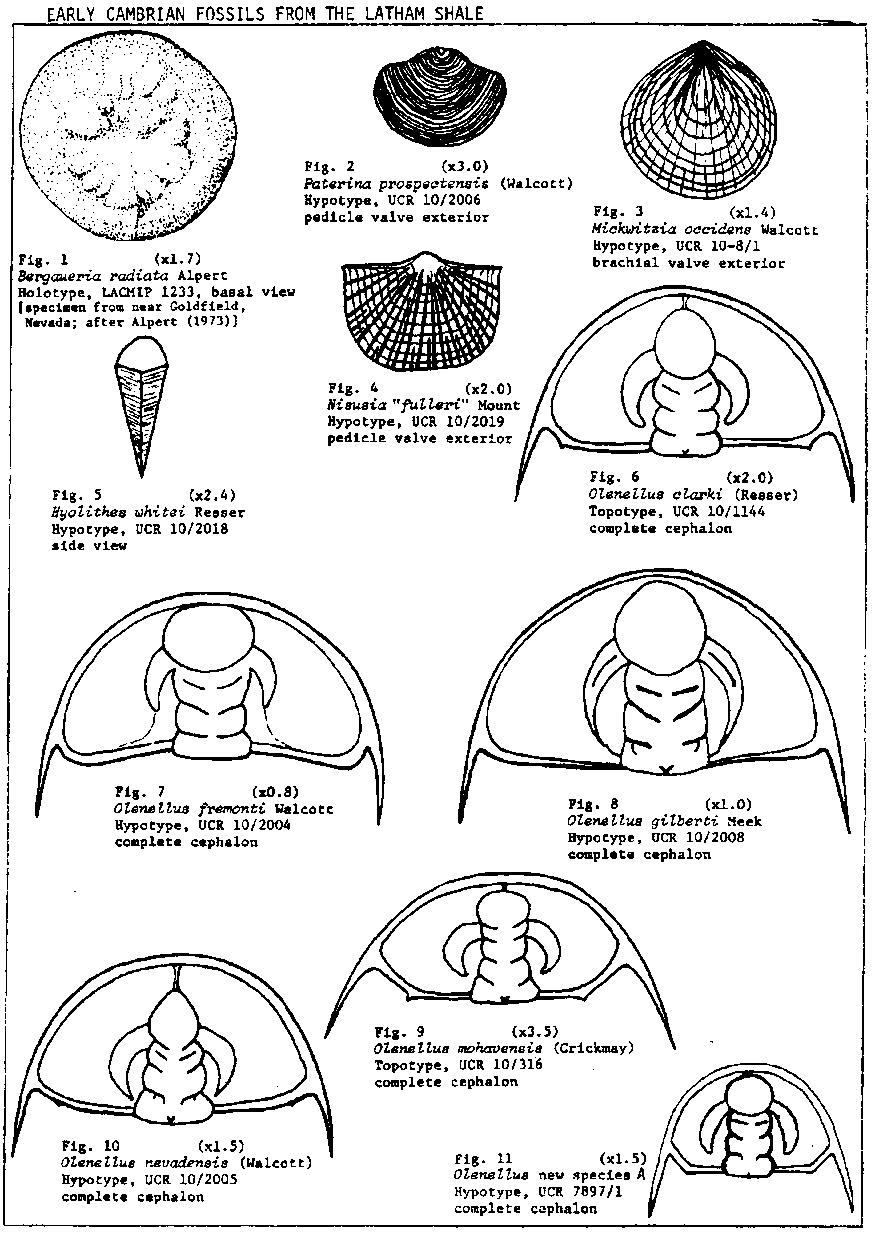
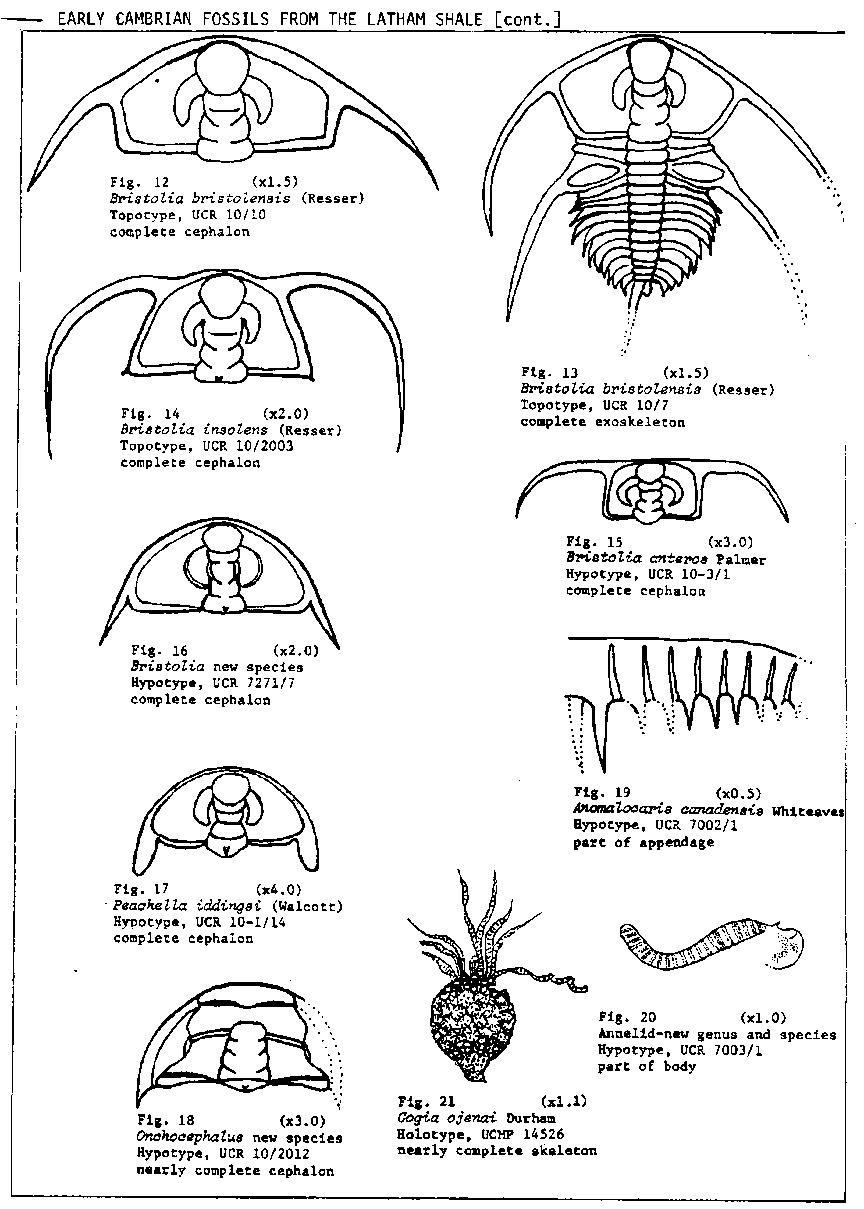

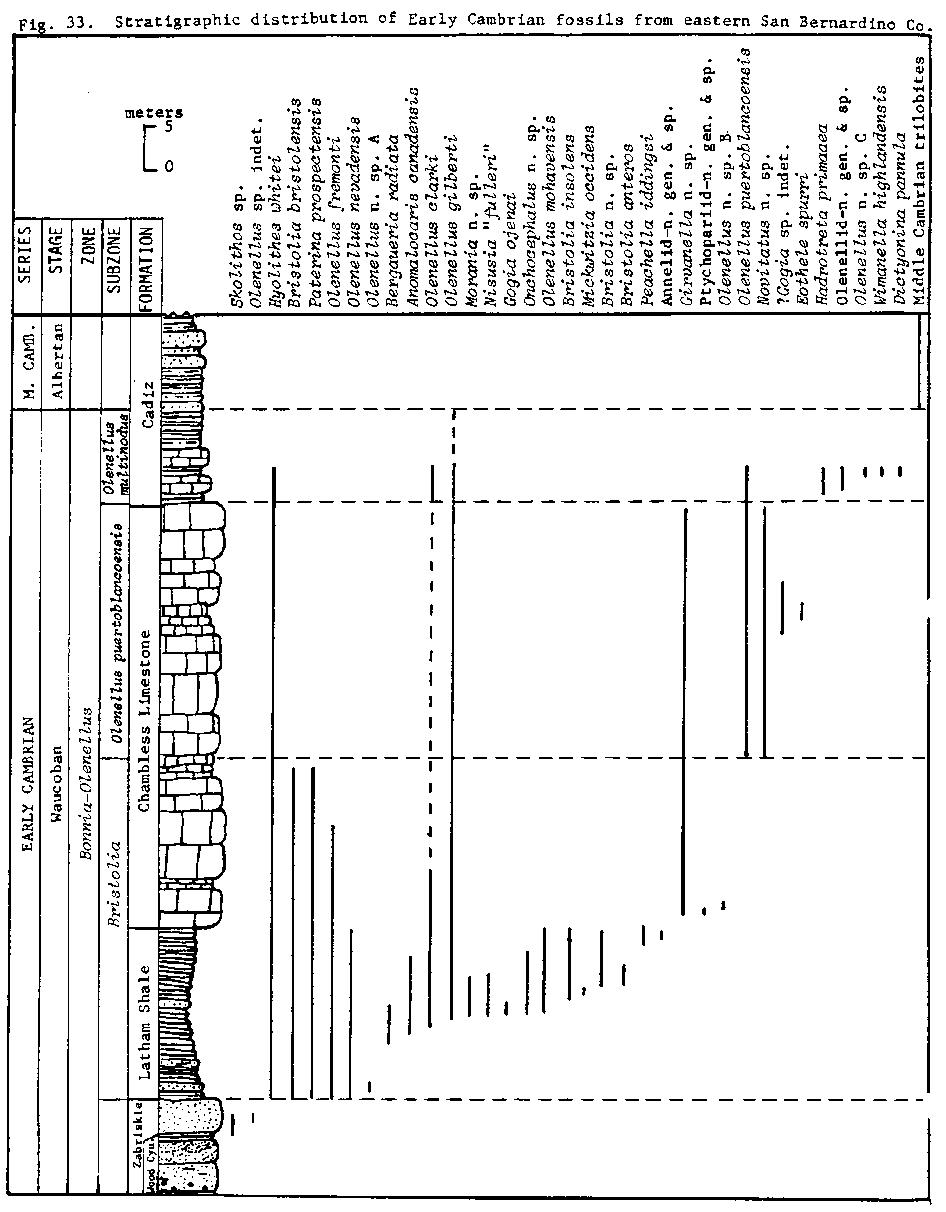
The oldest of the three subzones, the Bristolia Subzone of Mount (1974a, 1974b), is based on an assemblage of olenellid trilobites dominated by the genera Olenellus and Bristolia and the base of the subzone is placed at the first appearance of Bristolia bristolensis. The ranges of B. insolens, B. anteros, B. n. sp., Olenellus mohavensis, 0. nevadensis, 0. n. sp. A, Peachella iddingsi, Onchocephalus n. sp., and the new ptychopariid are apparently restricted to this subzone. The longer ranging Olenellus clarki and 0. gilberti appear to have their first occurrences within the subzone. The Bristolia Subzone ranges from the base of the Latham Shale to about 15 to 30 meters above the base of the Chambless Limestone.
The next subzone is the Olenellus puertoblancoensis Subzone of Mount (1976). The base of this unit is defined by the first occurrence of 0. puertoblancoensis. The upper boundary of this subzone approximates the Chambless Limestone-Cadiz Formation contact.
The youngest subzone of the Early Cambrian is the Olenellus multinodus Subzone. It is defined by the first occurrence of 0. multinodus Palmer which is found immediately above the base of the Cadiz Formation. Palmer and Halley (1979) first recognized this unit as their 0. multinodus Zonule. They reported the occurrence of this distinctive trilobite from the base of the Cadiz Formation at the type locality of the Chambless Limestone in the central Marble Mountains. It has not been collected during the present study. In the Marble Mountains the ranges of Olenellus n. sp. C and the new olenellid (see fig. 32) are restricted to this subzone. In the east-central California and Nevada area the assemblages from this subzone are charcterized by several species of Olenellus and the ptychopariid trilobites Crassifimbra, Zacanthopsis and Antagmus (Palmer, 1971; Palmer and Halley, 1979).
The three subzones are particularly well developed at the southern end of the Marble Mountains and on the west slope of the Providence Mountains just south of Summit Springs and these localities should serve as valuable reference sections for the subzones. Assemblages characteristic of these units have wide distribution in the southern Great Basin.
The middle Cambrian portion of the Cadiz Formation contains faunas from the Middle Cambrian Plagiura-Poliella (Hazzard, 1954), Albertella (Takeo Susuki, personal communication), and Glossopleura (Mason, 1935; Hazzard and Mason, 1936; Hazzard, 1954; Stoyanow and Susuki, 1955; Stoyanow, 1956; Fuller, 1980) Zones; however, they have not received critical biostratigraphical study.
Briggs, D. E. G., 1979. Anomalocaris, the largest known Cambrian arthropod. Palaeontology 22:631-664.
Clark, C. W., 1921. Lower and middle Cambrian formations of the Mohave Desert. Univ. California Publ. Bull. Dept. Geol. Sci. 13:1-7.
Cloud, P., 1973. Possible stratotype sequences for the basal Paleozoic in North America. American Jour. Sci. 273:193-206.
Darton, N. H., 1907. Discovery of Cambrian rocks in southeastern California. Jour. Geol. 15:470-475.
__________, 1915. Guidebook of the western United States: Part C, The Santa Fe route, with side trip to the Grand Canyon of the Colorado. U. S. Geol. Survey Bull. 613:149-153.
Durham, J. W., 1978. A lower Cambrian eocrinoid. Jour. Paleon. 52:195-199.
Fritz, W. H., 1972. Lower Cambrian trilobites from the Sekwi Formation type section, Mackenzie Mountains, northwestern Canada. Geol. Survey Canada Bull. 212:1-90.
__________, 1975. Broad correlations of some lower and middle Cambrian strata in the North American Cordillera. Geol. Survey Canada Paper 75-1(A):533-540.
Fuller, J., 1980. A Middle Cambrian fossil locality in the Cadiz Formation, Marble Mountains, California. in Paleontological tour of the Mojave Desert, California-Nevada. So. California Paleon. Soc. Spec. Publ. 2:30-34.
Hazzard, J. C., 1933. Notes on the Cambrian rocks of the eastern Mohave Desert, California. Univ. California Publ. Bull. Dept. Geol. Sci. 23:57-80.
__________, 1937a. Cambrian "Girvanella" from the southern Great Basin region (abstr.). Geol. Soc. America Proc. 1936 p. 354-355.
__________, 1937b. Paleozoic section in the Nopah and Resting Springs Mountains, Inyo County, California. California Jour. Mines Geol. 33:273-339.
__________, 1938. Paleozoic section in the Providence Mountains, San Bernardino County, California (abstr.). Geol. Soc. America Proc. 1937 p. 240-241.
__________, 1954. Rocks and structure of the northern Providence Mountains, San Bernardino County, California. California Div. Mines Bull. 170(4):27-35.
__________, and Mason, J. F., 1936. Middle Cambrian formations of the Providence and Marble Mountains, California. Geol. Soc. America Bull. 47:229-240.
Hewett, D. F., 1956. Geology and mineral resources of the Ivanpah Quadrangle, California and Nevada. U. S. Geol. Survey Prof. Paper 275:1-172.
Lanphere, M. A., 1964. Geochronologic studies in the eastern Mojave Desert, California. Jour. Geol. 72:381-399.
Mason, J. F., 1935. Fauna of the Cambrian Cadiz Formation, Marble Mountains, California. So. California Acad. Sci. Bull. 34:97-119.
Mount, J. D., 1973. Early Cambrian fauna of the Latham Shale, southern California (abstr.). So. California Acad. Sci. Ann. Mtg. Abstracts of Papers p. 10.
__________, 1974a. Early Cambrian faunas from the Marble and Providence Mountains, San Bernardino County, California. Bull. So. California Paleon. Soc. 6:1-5.
__________, 1974b. Biostratigraphy of the lower Cambrian in southeastern California (abstr.). Geol. Soc. America Abstracts With Programs 6(3):224.
__________, 1974c. Early Cambrian articulate brachiopods from the Marble Mountains, San Bernardino County, California. Bull. So. California Paleon. Soc. 6:47-52.
__________, 1976. Early Cambrian faunas from eastern San Bernardino County, California. Bull. So. California Paleon. Soc. 8:173-182.
__________, in press. A new articulate brachiopod from the lower Cambrian Latham Shale, southeastern California. So. California Acad. Sci. Bull. 79.
Nelson, C. A., 1976. Late Precambrian-Early Cambrian stratigraphic and faunal succession of eastern California and the Precambrian-Cambrian boundary. in Depositional environments of lower Paleozoic rocks in the White-Inyo Mountains, Inyo County, California. Pacific Coast Paleogeog. Field Guide 1:31-42.
Nolan, T. B., 1929. Notes on the stratigraphy and structure of the northwest portion of Spring Mountain, Nevada. American Jour. Sci. 17:461-472.
Palmer, A. R., 1971. The Cambrian of the Great Basin and adjacent areas, western United States. Cambrian of the New World, Wiley-Interscience, London, p. 1-78.
__________, and Halley, R. B., 1979. Physical stratigraphy and trilobite biostratigraphy of the Carrara Formation (Lower and middle Cambrian) in the southern Great Basin. U. S. Geol. Survey Prof. Paper 1047:1-131.
Rasetti, F., 1951. Middle Cambrian stratigraphy and faunas of the Canadian Rocky Mountains. Smithsonian Misc. Colln. 116(5):1-277.
Resser, C. E., 1928. Cambrian fossils from the Mohave Desert. Smithsonian Misc. Colln. 81(2):1-14.
Riccio, J. F. , 1949. Lower Cambrian fauna of the Marble Mountains, California. Compass of Sigma Gamma Epsilon 26:354-359.
__________, 1952. The lower Cambrian Olenellidae of the Marble Mountains, California. So. California Acad. Sci. Bull. 51:25-49.
Stewart, J. H., 1970. Upper Precambrian and lower Cambrian strata in the southern Great Basin, California and Nevada. U. S. Geol. Survey Prof. Paper 620:1-206.
Stoyanow, A., 1956. Types of Bathyuriscus howelli var. lodensis Clark. Geol. Soc. America Bull. 67:679-682.
__________, and Susuki, T., 1955. Discovery of Sonoraspis in southern California. Geol. Soc. America Bull. 66:467-470.
Sundberg. F. A., 1974. Distortion factor of Latham Shale trilobites. Bull. So. California Paleon. Soc. 6:121-124.
I particularly would like to thank the following individuals who collected and donated many interesting specimens pertinent to this study: James E. Fuller, Thomas Hill, John Kniffen, Alice M. Krueper, James D. L'Eclair, Gerald Licari, Mike Macomber, Robert F. Meade and Harold S. Meals. I also would like to thank Fred S. Sundberg and Takeo Susuki for their generous help.
__________
¹Department of Earth Sciences, University of California, Riverside, CA 92521.
![]() Return to Paleontology Resources page
Return to Paleontology Resources page
![]() Return to first page of Jack's Home Place
Return to first page of Jack's Home Place
|
|
|
 | |
California NetRing | |